INTRODUCTION
The mitochondrial life cycle (e.g., mitochondrial biogenesis, mitochondrial dynamics, and mitochondrial removal by mitophagy) is carefully orchestrated to ensure the efficient generation of cellular energy. These processes maintain reactive oxygen species (ROS) production within an optimal range for cellular health. Defective cellular energy metabolism has been suggested as a significant cause of metabolic and age-related diseases, and obesity, diabetes, and aging share mitochondrial impairment as a common pathophysiological mechanism 1–4. Moreover, mitochondrial dysfunction has been shown to underlie several cardiac diseases; it is well-known that the energetic failure, which characterizes the failing heart, is mainly due to a mitochondrial energy deficiency 5, 6. Therefore, activation of mitochondrial biogenesis is paramount for the prevention and treatment of energy metabolism and age-related diseases.
Calorie restriction (CR), a low-calorie dietary regimen without malnutrition, decreases the incidence of several age-associated disorders and is considered the gold-standard, nongenetic approach for life span extension 7. A growing body of evidence in several organisms demonstrates that an increase in mitochondrial activity, together with activation of the ROS defense system, is associated with the health effects of the CR regimen 7, 8.
Previous observations in yeast suggested that the branched-chain amino acids (BCAAs), including leucine, isoleucine, and valine, might be potential candidates in promoting life span extension 9. We have demonstrated that long-term dietary supplementation with a specific BCAA-enriched amino acid mixture (BCAAem) increased the average life span of male mice 10. This effect was accompanied by increased mitochondrial biogenesis and sirtuin 1 (SIRT1) expression, and an upregulated ROS defense system, with reduced oxidative damage, both in the cardiac and skeletal muscles of middle-aged mice 10. We confirmed the efficacy of the BCAAem mixture in different pathologies: supplementation of this mixture prevented muscular damage induced by rosuvastatin through induction of de novo protein synthesis and preservation of mitochondrial efficiency , ameliorating the control of oxidative stress in the statin-treated mice and muscle cells 11. Moreover, we observed that dietary supplementation of the BCAAem mixture prevented fat accumulation and mitochondrial dysfunction in hepatocytes of alcohol-consuming rats 12.
Our present, preliminary study aimed to identify novel amino acid (AA) mixtures capable of promoting mitochondrial function with an efficiency higher than that of classic BCAAem formula. To this aim, we investigated a combination with the same stoichiometric AA ratio of BCAAem (Leu: Iso: Val 2:1:1), E7, and another formula of an AA mixture with a stoichiometric AA ratio different from BCAAem (3:1:1 Leu: Iso: Val vs. 2:1:1, as in BCAAem), α5. Furthermore, both mixtures were enriched with the tricarboxylic acid cycle (TCA) substrates, including citrate, succinate, and malate (ratio 8:2:2) (Table 1).
TABLE 1. Composition of the Mixtures.
| Mixture | BCAAem (% ) |
α5 (%) |
E7 (%) |
|---|---|---|---|
| L-Leucine | 30.01 | 31.09 | 22.45 |
| L-Lysine (chlorhydrate) | 19.58 | 16.90 | 21.13 |
| L-Isoleucine | 15.00 | 10.36 | 11.23 |
| L-Valine | 15.00 | 10.36 | 11.23 |
| L-Threonine | 8.40 | 7.25 | 13.1 |
| L-Cysteine | 3.60 | 3.11 | 2.81 |
| L-Histidine | 3.60 | 3.11 | 2.81 |
| L-Phenylalanine | 2.40 | 2.07 | 1.87 |
| L-Methionine | 1.20 | 1.04 | 0.94 |
| L-Tyrosine | 0.72 | 0.62 | – |
| L-Tryprophan | 0.48 | 2.07 | 0.94 |
| Vitamin B1 (thiamine chlorhydrate) | – | 0.004 | 0.02 |
| Vitamin B6 (piridoxine chlorhydrate) | – | 0.004 | 0.02 |
| Citric acid | – | 8.00 | 7.65 |
| Malic acid | – | 2.00 | 1.92 |
| Succinic acid | – | 2.00 | 1.92 |
| Ratio Leucine: Isoleucine:Valine | 2:1:1 | 3:1:1 | 2:1:1 |
BCAAem, branched-chain amino acid enriched mixture.
HL-1 cells are beating cardiomyocytes that display spontaneous contraction in vitro and, as such, represent an excellent model of cardiac function 13. As heart energy metabolism strongly relies on mitochondrial substrate oxidation, these mitochondria-rich cells can be suitably used to verify the efficacy of the new AA mixtures on mitochondrial function. To this aim, we have evaluated the gene expression of the main mitochondrial biogenesis and function markers, peroxisome proliferator-receptor γ coactivator 1 α (PGC-1α), mitochondrial DNA (mtDNA), transcription factor A (Tfam), and cytochrome c (Cyt c), as well as mtDNA amount. Moreover, we have analyzed the expression of genes involved in BCAA catabolism and, to confirm increased mitochondrial function, we have evaluated the oxygen consumption rate (OCR) in HL-1 cardiomyocytes exposed to these novel AA mixtures. We hypothesized that adding TCA substrates could improve the mitochondrial oxidative capacity of the selective AA mixtures.
MATERIALS AND METHODS
Cells and treatments
HL-1 cardiomyocytes (a gift from W.C. Claycomb) were plated in fibronectin-/gelatin-coated flasks, grown to 70–80% confluence in Claycomb medium (JRH Biosciences) supplemented with 100 μM norepinephrine (from a 10 mM norepinephrine [Sigma-Aldrich] stock solution dissolved in 30 mM L-ascorbic acid [Sigma-Aldrich]), 2 mM L-glutamine, 100 U/mL penicillin, 100 μg/mL streptomycin, and 10% fetal bovine serum (FBS, JRH Biosciences). Cells were treated with 1% of the mixtures (composition is reported in Table 1) for 48 h. Untreated cells were plated as controls. Every 24 h, media were replaced in both control and treatment flasks, with fresh media, with AA mixtures. At the end of the experimental treatments, cells were used for mRNA or DNA extraction, or for assessment of oxygen consumption.
Total RNA and DNA extraction, and gene expression analysis
RNA was isolated from HL-1 cardiomyocytes using the RNeasy Mini Kit (Qiagen), and cDNA was synthesized using the iScript cDNA Synthesis Kit (Bio-Rad Laboratories). The gene expression (primer sequences in Table 2) relative level was calculated as 2-DDCT, in which DDCT corresponded to the difference between the DCT of either treatment and the DCT of the untreated group.
TABLE 2. Primers for Quantitative Real Time-Polymerase Chain Reaction.
| Primer Sense (5'-3') | Primer Antisense (5'-3') | Ta (°C) | |
|---|---|---|---|
| GAPDH | AACTTTGGCATTGTGGAAGG | ACACATTGGGGGTAGGAACA | 60 |
| Cyt c | ATAGGGGCATGTCACCTCAAAC | GTGGTTAGCCATGACCTGAAAG | 61 |
| PGC1-α | ACTATGAATCAAGCCACTACAGAC | TTCATCCCTCTTGAGCCTTTCG | 61 |
| Tfam | AAGACCTCGTTCAGCATATAACATT | TTTTCCAAGCCTCATTTACAAGC | 60 |
| KFL15 | ACACCAAGAGCAGCCACCTCA | TGAGATCGCCGGTGCCTTGA | 60 |
| PP2CM | ACCACAGGCAGGCGACTC | TGGCTCATCAATGCGGTTATCC | 60 |
| mtDNA | ACATGCAAACCTCCATAGACCGG | TCACTGCTGAGTCCCGTGGG | 63 |
| gDNA | GGTCGCGGTGTGGGCATTTG | CGTGATCGTAGCGTCTGGTT | 60 |
GAPDH, glyceraldehyde 3-phosphate dehydrogenase; Cyt c, cytochrome c; PGC1-α, peroxisome proliferator-receptor γ coactivator 1 α; Tfam, transcription factor A; KLF15, Krüppel-like factor 15; PP2CM, protein phosphatase 2 Cm; mtDNA, mitochondrial DNA; gDNA, genomic DNA (chromosome 5 Actb).
For mtDNA analysis, total DNA was extracted with a QIAamp DNA extraction kit (QIAGEN). mtDNA was amplified using primers specific for the D-Loop region of mouse mtDNA gene and normalized to the levels of the intronic region of the GAPDH gene (gDNA). Primers were designed using Beacon Designer 2.6 software (Premier Biosoft International; Palo Alto, CA).
Oxygen consumption
OCR in HL-1 cells was measured with The Seahorse XFe24 Analyzer (Agilent Technologies, Santa Clara, CA, USA) 14. Briefly, 1 × 105 cells per well, cultured in their complete media, were plated in a 96 well Seahorse plate. Then, cells were washed twice with Seahorse assay media (Agilent Technologies, Santa Clara, CA, USA) and incubated for 30 min in a CO2-free incubator at 37°C. Cell respiration was determined using the Mito Stress Test kit (Agilent Technologies, Santa Clara, CA, USA) according to the manufacturer’s instructions. After basal OCR, oligomycin (2 μM), FCCP (Carbonyl cyanide 4-(trifluoromethoxy) phenylhydrazone) (1.5 μM), Rotenone (0.5 μM), and Antimycin (0.5 μM) were sequentially added in the assay to assess uncoupled, maximal, and nonmitochondrial OCR, respectively. After the test, total proteins were extracted with Ripa cell lysis and extraction buffer (Thermo Scientific), and protein content was estimated using BCA protein assay kit (Pierce Thermo Scientific); OCR was then normalized with total cellular protein amount value, as recommended by the manufacturer using Wave 2.0 desktop software. Normalized data were further analyzed for interpretation using the Seahorse XF Cell Mito stress test Report generator.
RESULTS
E7 and α5 mixtures are more effective than BCAAem in inducing mitochondrial biogenesis
As described in Table 1, the E7 mixture’s composition differs from BCAAem (the classic AA mixture). Although the Leucine:Isoleucine:Valine ratio of E7 is the same as the BCAAem mixture (2:1:1), E7 is enriched with the TCA cycle substrates, namely, citric acid, succinic acid, and malic acid. Moreover, E7 does not contain L-tyrosine and has a higher concentration of L-lysine than BCAAem. The α5 mixture, together with citric acid, succinic acid, and malic acid, also includes vitamins B1 and B6. Furthermore, the stoichiometric BCAA ratio of α5 (3:1:1) is different from the BCAAem mixture.
To compare the efficacy of the new AA formulas, we first analyzed the mtDNA amount in HL-1 cells treated with 1% of the AA mixtures for 48 h. As shown in Figure 1, as expected, the BCAAem mixture significantly increased the mtDNA amount. However, both α5 and E7 were more effective than BCAAem in increasing mtDNA levels (+15% for α5 and +16% for E7 vs. BCAAem), indicating a stronger stimulation of mitochondrial biogenesis.
FIG 1. mtDNA levels in HL-1 cardiomyocytes left untreated (CT) or incubated with the three different mixtures 1% for 48 h. Quantitative PCR was performed in triplicate and normalized to gDNA (n = 3, mean ± SEM). *P < 0.05 versus untreated cells, expressed as 1.0. **P < 0.01 versus untreated cells. #P < 0.05 versus BCAAem.

We further investigated the effects of the mixtures on mitochondrial biogenesis in HL-1 cells by assessing the gene expression of mitochondrial biogenesis markers. After 48 h of supplementation, all AA mixtures significantly increased the mRNAs levels of PGC-1a, Tfam, and Cyt c (Figure 2) than controls. However, and in line with mtDNA data, α5 and E7 were more effective than BCAAem. When compared to BCAAem, both mixtures led to a higher increase in mRNA levels of all the mitochondrial biogenesis markers. PGC-1α, Tfam, and CytC were increased by 24%, 32%, and 15% more by α5, and 40%, 33%, and 20% more by E7 (Figure 2).
FIG 2. Mitochondrial biogenesis markers in HL-1 cardiomyocytes treated with 1% mixtures for 48 h. Quantitative PCR, performed in triplicate and normalized to GAPDH of PGC1α (left panel), CytC (central panel), and Tfam (right panel) (n = 3, mean ± SEM). *P < 0.05 versus untreated cells, expressed as 1.0. #P < 0.05 versus BCAAem.

These results suggest that the novel formulas, enriched with the TCA cycle substrates, including citric acid, succinic acid, and malic acid, are more potent inducers of mitochondrial biogenesis than BCAAem formula.
Both α5 and E7 show greater efficiency in activating the BCAA catabolic pathway than BCAAem
It is well-known that the entire BCAA catabolic pathway is located within the mitochondrial matrix. The first steps in BCAA catabolism are common to the three BCAAs and require the mitochondrial isoform of BCAA aminotransferase (BCATm) and branched-chain α-keto acid dehydrogenase complex (BCKDC). In the first and fully reversible step of degradation, mitochondrial BCAT transfers the amino group from BCAAs to a-ketoglutarate to form the corresponding branched-chain a-keto acids (BCKAs) and glutamate. After that, BCKDC catalyzes the decarboxylation of the carboxyl groups of BCKAs, to create the corresponding branched-chain acyl-CoA esters. This reaction is irreversible and, therefore, commits the BCAAs to degradation. Besides end-product allosteric inhibition by nicotinamide adenine dinucleotide reduced form (NADH), α-ketoisocaproate, and branched-chain acyl-CoA esters, BCKDC activity is regulated by phosphorylation regulatory subunit E1a by the BCKDC kinase BDK, which leads to the inhibition of BCKDC activity.
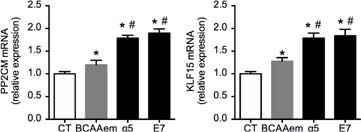
On the other hand, E1a is dephosphorylated by the mitochondrial Mg2+/Mn2+ dependent Ser/Thr protein phosphatase PP2Cm, leading to BCKDC activation 15. Moreover, the transcription factor KLF15 has been shown to increase the cardiac gene expression of BCAT, BCKDC, and PP2Cm 16. To investigate if the high mitochondrial biogenetic capacity of α5 and E7 was associated with the activation of the BCAA catabolic pathway, we assessed the mRNA levels of PP2Cm and KLF15 in HL-1 cells treated with the three mixtures for 48 h.
As shown in Figure 3, each formula increased PP2Cm and KLF15 mRNA levels for controls in HL-1 cardiomyocytes. However, α5 and E7 increased both mRNAs to a greater extent compared to BCAAem. These data indicate that, although all the three mixtures can recruit and activate the BCAA catabolic pathway, α5 and E7 are more effective than BCAAem.
FIG 3. PP2CM and KFL15 mRNA in HL-1 cardiomyocytes treated with 1% mixtures for 48 h. Quantitative PCR, performed in triplicate and normalized to GAPDH of PP2CM (left panel) and KLF15 (right panel) (n = 3, mean ± SEM). *P < 0.05 versus untreated cells, expressed as 1.0; #P < 0.05 versus BCAAem.
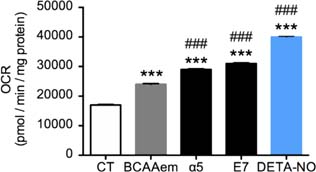
Mitochondrial oxygen consumption is higher in HL-1 cardiomyocytes supplemented with α5 and E7
To further confirm the efficacy of these new formulas on mitochondrial function, we investigated their ability to promote oxygen consumption. As a positive control and reference, we treated cells with well-known mitochondrial activator nitric oxide (NO) 17. Indeed, NO was found to trigger mitochondrial biogenesis in cells as diverse as brown adipocytes and 3T3-L1, U937, and HeLa cells. This effect of NO was dependent on cyclic guanosine 3’,5’-monophosphate (cGMP), and was mediated by the induction of PGC-1α, a master regulator of mitochondrial biogenesis 17. We exposed the HL-1 cells to either E7 and α5 mixtures (1%) or 50 µM of the NO-donor DETA-NO, known to increase oxygen consumption 17. As expected, after 48 h of DETA-NO treatment, we observed a rise in the OCR (Figure 4). However, while BCAAem increased OCR as well, both α5 and E7 were more effective than BCAAem in stimulating OCR (+25% [α5] and +30% [E7] vs. BCAAem) (Figure 4). These data are in line with those on mtDNA and mitochondrial gene expression, confirming the ability of a5 and E7 to stimulate mitochondrial function to a greater extent than BCAAem.
FIG 4. Oxygen consumption rate in HL-1 cardiomyocytes treated with 1% mixtures or DETA-NO for 48 h. ***P < 0.001 versus untreated cell; ###P < 0.001 versus BCAAem. Samples are n = 4 and mean ± SEM.
DISCUSSION
Overall, our preliminary results suggest that E7 and α5 (1%) are significantly more active than classical BCAAem formula in promoting mitochondrial biogenesis, mitochondrial function, and BCAA catabolism in HL-1 cardiomyocytes. Although E7 and α5 contain a lower percentage of BCAA than BCAAem (Table 1), their effect on the expression of BCAA catabolic genes is higher (Figure 3). These data suggest that, besides the absolute amount of the three BCAAs and their ratios, the combination of these two factors is probably a more critical issue in the regulation of BCAA pathway activation, in line with previous reports 18, 19.
Given the evidence on relevant health beneficial effects observed with the previous BCAAem mixture, multiple potential clinical implications for the new formulas may be hypothesized. BCAAem was found to reduce sarcopenia in elderly subjects 20, improve exercise capacities in elderly subjects affected by chronic heart failure 21, markedly reduce the incidence of infections acquired in geriatric, long-term rehabilitation centres 22, increase the serum albumin and total proteins in hemodialysis patients, with reduction of inflammation markers and correction of anaemia 23, in addition to improving gas exchange and cognitive function score in patients with severe chronic obstructive pulmonary disease 24. Most importantly, we recently reported that 2 months of BCAAem supplementation improves not only nutritional status, general health, and muscle mass but also cognitive performance in elderly, malnourished patients 25. These effects are positively associated to increased adenosine triphosphate (ATP) production, mitochondrial respiration, and low oxidative stress in peripheral blood mononuclear cells 25. Moreover, the activation of BCAA catabolism in supplemented cardiomyocytes indicates that these original mixtures could serve as an innovative nutritional approach for the management of heart failure, in addition to the well-established pharmacological approaches 16.
Several hypotheses can be put forward to explain the mechanisms of these new formulas on mitochondrial activity. A possibile suggestion is that the supply of citrate, malate, and succinate could enhance mitochondrial oxidative capacity by merely providing more substrates to the TCA cycle. For example, the amount of citrate, as the critical first substrate of the TCA cycle, could enhance the aconitase activity. Moreover, the catabolism of BCAA, AAs enriched in the mixture, and acetyl-CoA also provide succinyl-CoA. This later could activate succinyl-CoA synthetase reaction, which, in turn, produces succinate as a substrate for the subsequent response from succinate dehydrogenase of the Krebs cycle. Providing succinate, along with BCAA, in the mixture could also stimulate the succinate dehydrogenase reaction, further boosting the Krebs cycle. Noteworthy, by directly providing FADH2, succinate dehydrogenase also becomes part of the mitochondrial electron transport chain (complex II). Its stimulation by succinate could instantly activate mitochondrial redox carriers and increase membrane potential, thus enhancing proton gradient, oxygen consumption, and ATP synthesis. At the same time, malate supplement could enable malate dehydrogenase reaction and increase NADH levels; this would also provide substrates for complex I and therefore increase ATP levels in the same manner as succinate-derived FADH.
On the other hand, malate could stimulate the activity of the malate–aspartate shuttle. This process would favor the entry of cytosolic NADH into mitochondria, which would otherwise be impermeable through mitochondrial membrane, thus rendering it available for mitochondrial oxidation. This biochemical step would further increase mitochondrial activity and oxygen consumption (Figure 4).
In this context, it is also noteworthy that, in α5 and E7 supplemented cells, the activation of BCAA catabolism would result in the continuous consumption of α-ketoglutarate from the TCA cycle through BCATm reaction, thus “draining off” this intermediate. On the other hand, the simultaneous supplementation of succinate from the mixture would feed the cycle by bypassing α-ketoglutarate (α-KG) dehydrogenase reaction and entering directly at the succinate dehydrogenase step. Thus, the activation of BCAA catabolism could be a cataplerotic reaction capable of preventing excessive α-KG accumulation. This later could derive from the continuous and massive TCA substrate supplementation from the mixtures in the mitochondrial matrix, and it could otherwise slow the cycle by exerting an inhibitory “back-pressure” 26. Therefore, the simultaneous activation of BCAA catabolism and TCA cycle would allow a constant, unrestrained flow of substrates and energy production.
The oxygen consumption measured with the Seahorse analyzer as a direct evaluation of the mitochondrial function may be considered a major strength of our study. However, we have reported only in vitro results. Further in vivo studies will be necessary to confirm the efficacy of these new AA formulas in promoting mitochondrial activity. In particular, given the essential role of mitochondria in energy metabolism, as well as in the aging process, it will be of great interest to verify their efficacy in trained compared to sedentary mice, in addition to aged compared to young animals. Translating into clinical practice, these in vitro and, hopefully, in vivo results are not obvious: the clinical trials are expensive, and the nutritional studies are very complex to organize and perform. Nevertheless, the accumulated evidence on the clinical efficacy of our previous AA mixture is very encouraging.
CONCLUSIONS
Our data suggest that E7 and α5 are significantly more active than BCAAem formula in promoting mitochondrial biogenesis, mitochondrial function, and BCAA catabolism in HL-1 cardiomyocytes. Further studies in rodents and humans are necessary to confirm the efficacy of these novel AA formulas to promote mitochondrial activity in vivo. Interestingly, a recent report shows that the administration of succinate can stimulate mitochondria-dependent thermogenesis in brown adipose tissue (BAT), leading to protection against diet-induced obesity and a glucose tolerance improvement 27. Furthermore, the thermogenesis process in BAT strongly relies on BCAA utlization and catabolism 28. Therefore, a healthy relationship and crosstalk exist between TCA cycle intermediates and BCAA catabolism, whose alteration underlies diseases linked to energy metabolism. Activation of both pathways, employing α5 and E7 mixtures, could greatly help in the prevention and treatment of these diseases using a novel nutritional approach.